Брусок черепной: описание, назначение, размеры, правила монтажа, техника выполнения работ и советы специалистов
описание, назначение, размеры, правила монтажа, техника выполнения работ и советы специалистов
Любое строительство отнимает много времени и сил. Особенно если стройка ведется своими руками. Нужно много знать: понимать принципы строительства объекта, свойства материалов, способы соединения конструкций. Нужны не только знания, но и опыт. Опытные строители работают быстро и качественно, могут отличить качественный материал от дешевой подделки, имеют все необходимые навыки стыковки и соединения конструкций. Новички в строительстве лишены всего этого. Все, что им остается — это собирать информацию и учиться на собственных ошибках. В этой статье будет рассказано о том, что такое черепные бруски, так в народе называют опорный брус, для чего они служат и как правильно смонтировать их на балке.
Назначение опорных брусков
Опорные бруски служат для крепления досок пола, потолка или стропильной системы. В старые времена, когда для балок применялся не брус, а цельное бревно, череп выбирался мастером с помощью топора и тесла. Сейчас некоторые строители в целях экономии тоже используют бревна для лаг, но череп пропиливают бензопилой, а затем выбирают топором или стамеской. В последнее время для лаг пола или потолка используют брус, а для крепления перекрытия на него набивают черепные бруски. Это связано с тем, что обычные бревенчатые дома сейчас редкость, и кругляк уже почти не продают, а на строительных базах брус разного сечения всегда есть в наличии.
Сейчас некоторые строители в целях экономии тоже используют бревна для лаг, но череп пропиливают бензопилой, а затем выбирают топором или стамеской. В последнее время для лаг пола или потолка используют брус, а для крепления перекрытия на него набивают черепные бруски. Это связано с тем, что обычные бревенчатые дома сейчас редкость, и кругляк уже почти не продают, а на строительных базах брус разного сечения всегда есть в наличии.
Способы монтажа чернового пола
Балки с черепными брусками в основном используют для настила чернового пола. Он, в свою очередь, служит для утепления перекрытия. В качестве чернового пола могут использоваться доски или листы OSB. Они укладываются на бруски, а поверх ложится утеплитель. Все балки, бруски и доски необходимо обработать антисептиком от жучка и плесени. Иногда черепной брусок используют для дополнительного крепления самих балок. В этом случае они набиваются по периметру нижней обвязки, а балки ложатся на них. Бруски для чернового пола можно закрепить поперек балок, прибивая их снизу.
Способы монтажа стропильных систем
Для монтажа стропильной системы тоже используется черепной брусок. Для чего он нужен? Он упрощает монтаж стропил, особенно сложных — вальмовых или яндовых крыш, там, где требуются перекрестные соединения и разной длины стропила. Для равномерного распределения нагрузки по основным несущим стропилам бруски используют как дополнительные опоры.
Монтаж проводится следующим образом. На хребет вальмы устанавливаются накосные стропила. К ним крепятся нарожники длинными саморезами или гвоздями. Но для хорошей прочности к соединениям необходимо между нарожниками с обеих сторон прибить черепной брусок, чтобы он своими концами распирал нарожники по всей длине накосных стропил. Это поможет равномерно распределить нагрузку по всем стропилам и повысит прочность всей конструкции. Брусок необходимо запилить под соответствующим углом ножовкой или торцовочной пилой. Таким же способом можно усилить стойки стропил.
Черепной брусок для крепления потолка
Для крепления потолка бруски можно использовать двумя способами, как и в случае с полом. Первый — это прибить бруски наравне с торцом балки. В этом случае черновой пол второго этажа или мансарды настилается сверху на бруски, а обрешетка потолка крепится снизу поперек балок, и уже на обрешетку монтируется чистовой вариант потолка. Во-втором варианте бруски пришиваются снизу поперек балок, а обрешетка крепится непосредственно на них. В том и другом случае можно использовать уже готовые балки с двумя черепными брусками. В дальнейшем между балками второго этажа прокладывают пароизоляцию и слой утеплителя.
Первый — это прибить бруски наравне с торцом балки. В этом случае черновой пол второго этажа или мансарды настилается сверху на бруски, а обрешетка потолка крепится снизу поперек балок, и уже на обрешетку монтируется чистовой вариант потолка. Во-втором варианте бруски пришиваются снизу поперек балок, а обрешетка крепится непосредственно на них. В том и другом случае можно использовать уже готовые балки с двумя черепными брусками. В дальнейшем между балками второго этажа прокладывают пароизоляцию и слой утеплителя.
Как крепить брусок к балке: советы специалистов
Крепить бруски к балке можно гвоздями или саморезами. Длина их подбирается в зависимости от толщины бруска. Чем он толще, тем длиннее гвоздь. Допускается крепление с двух сторон: со стороны бруска или со стороны балки. Можно прибить, чередуя оба способа. Важно соблюдать ровную плоскость, чтобы торцы бруска были наравне с балкой.
Крепить брусок нужно через одинаковые отрезки, визуально разделив расстояние между его концами. Если вы пользуетесь гвоздями, то уделите внимание молотку. Во избежание травм инструмент выбирают с нескользящей рукояткой и ровным, плоским бойком. Если боек избит, то молоток может соскальзывать со шляпки гвоздя, тем самым травмируя руку. Черепные бруски выбирают ровные, со строганой поверхностью. Забивать гвоздь нужно до самой шляпки, делая два-три контрольных удара для более плотного сопряжения поверхностей.
Если вы пользуетесь гвоздями, то уделите внимание молотку. Во избежание травм инструмент выбирают с нескользящей рукояткой и ровным, плоским бойком. Если боек избит, то молоток может соскальзывать со шляпки гвоздя, тем самым травмируя руку. Черепные бруски выбирают ровные, со строганой поверхностью. Забивать гвоздь нужно до самой шляпки, делая два-три контрольных удара для более плотного сопряжения поверхностей.
Для крепления бруска саморезами применяется шуруповерт с битой. Саморезы применяются с широким шагом резьбы и защитным покрытием. Оксидированные саморезы при контакте с агрессивной средой быстро ржавеют, поэтому, чтобы закрепить черновые полы, черепной брусок должен быть прикручен на желтые или белые саморезы. При закручивании важно уделять внимание состоянию биты. Грани ее должны быть целыми, если грани стерты, то бита будет проскакивать в шляпке самореза, не обеспечивая плотный контакт между поверхностями. На трещотке шуруповерта нужно подобрать такое значение, при котором обеспечивается наиболее плотное примыкание поверхностей.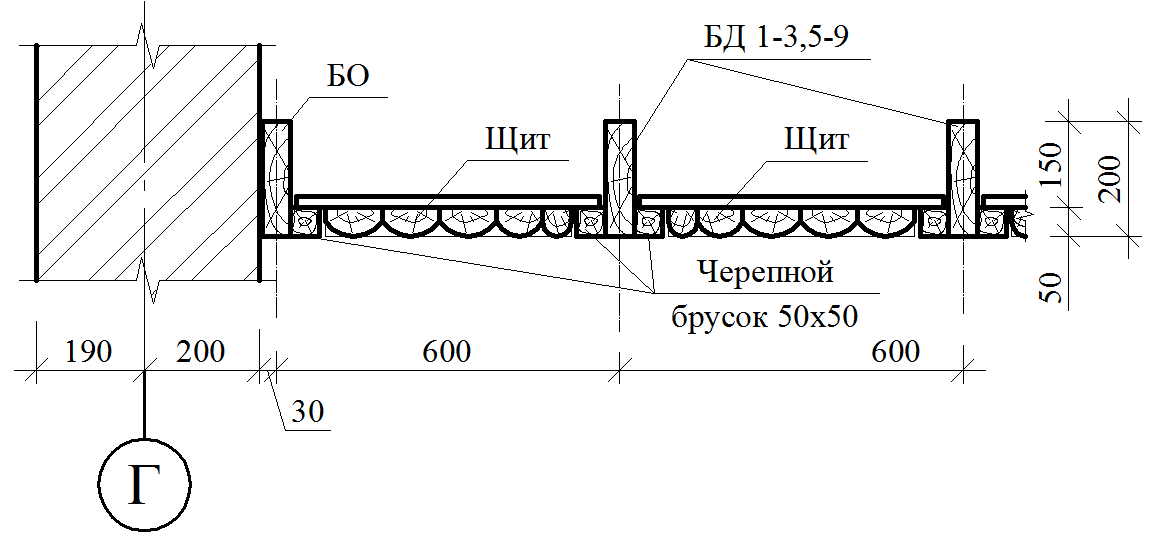
Размеры опорных брусков
Самый главный недостаток при использовании опорных брусков — это уменьшение полезного объема для утеплителя. Чем больше брусок, тем меньше утеплителя можно положить.
Стандартные размеры черепного бруска — 50 x 50 мм. Некоторые мастера, для того чтобы увеличить объем для утеплителя, используют размеры 40 x 50 мм или даже 20 x 40 мм. Но такая экономия может выйти боком. Меньшая толщина бруса имеет сниженный коэффициент надежности такого крепления. Для того чтобы воспользоваться узкими брусками, нужно уменьшить расстояние между балками. Значит, при расчетах придется добавлять одну или две балки. Это не очень экономно, так как черепной брус намного дешевле толстой балки. Лучше всего придерживаться стандартных размеров, рассчитанных на большую нагрузку.
В заключение
Стройка дома — всегда ответственное испытание для хозяина. Знать, что такое черепные бруски и как их установить, — это только один из моментов, который необходимо запомнить домашнему умельцу. На самом деле этапов строительства много, от правильной заливки фундамента до крепления конька. Но если уже удалось возвести дом под крышу, это не значит, что тотчас можно переехать туда жить.
На самом деле этапов строительства много, от правильной заливки фундамента до крепления конька. Но если уже удалось возвести дом под крышу, это не значит, что тотчас можно переехать туда жить.
Внутренняя отделка может быть более затратной, чем сам каркас. Дом нужно утеплить, обшить или оштукатурить стены, сделать отопление, обустроить канализацию, провести водопровод, газ. После завершения строительства дома останется еще тысяча незавершенных дел: уборка мусора, благоустройство участка, постройка бани, подсобных помещений. Но даже когда все будет завершено, спокойная жизнь не гарантирована. В своем доме всегда найдутся дела, которые потребуют дальнейшего вмешательства.
Подшивка перекрытий и установка утеплителя. Альтернатива черепным брускам.
|
Прежде всего необходимо понять, каки функции должна выполнять подшивка балок перекрытия: Традиционная подшивка на черепных брусках неплохо справляется с функциями поддержки утеплителя и условно хорошо с защитой от ветра и грызунов. (Злоумышленников мы оставим — это все-таки экзотика). А вот с функциями транспорта водяного пара и придания дополнительной механической прочности деревянная подшивка на черепных брусках не справляется. Доски из доски дюймовки обладают в 10 раз более низкой паропроницаемостью поперек волокон, по сравнению с минеральной ватой утеплителя. Свободная же укладка черепных досок на черепные бруски и даже их подшивка (получается двойной шарнир) не обеспечивают создания единой горизонтальной связи между балками перекрытия. И, следовательно, не увеличивают прочности всей конструкции в целом. Самый же главный недостаток технологии подшивки перекрытия по черепным брускам — это уменьшение эффективной толщины утеплителя в перекрытию. Еще один вариант решения в данном случае — это использование ЭППС в качестве утеплителя. И то вам понадобиться 12-13 см ЭППС в Москве или Петербурге… Обычный пенопласт требует той же толщины, что и минераловатный утеплитель, а резол-формальдегидный пенопласт — даже большей, чем минеральная вата (он обладает высокой теплопроводностью при работе во влажных условиях). |
|
|
Как же обойтись без устаревших черепных брусков в подшивке балок перекрытий? Наиболее рациональной является нижняя подшивка балок перекрытий. Нижняя подшивка балок перекрытий позволяет использовать всю высоту балки для размещения утеплителя. Нижняя подшивка досками или OSB-3 позволяет создать прочные горизонтальные связи между балками увеличивающие несущую способность перекрытия в целом. |
|
|
Возможна комбинация обрешетки и стальной сетки — для защиты от проникновения грызунов в перекрытие. Для влаго-ветрозащиты утеплителя и перекрытия в случае использования обершетки или сетки, лучше всего использовать трехслойные прочные супердиффузионные мембраны типа Изоспан AS или Мегаизол SD. Обычно эти мембраны применяются в подкровельном пространстве. В отличие от более дешевого Изоспана А, эти мембраны обладают атмосферостойкостью, большей долговечностью и не требуют зазора между собой и утеплителем. В случае экономии можно использовать и Изоспан А. |
|
|
Оптимальная схема монтажа нижней подшивки выглядит так: снизу под балки степлером подшивается с нахлестом и их проклейкой двусторонним скотчем супердиффузионная мембрана, затем с помощью гвоздей или 25 мм скоб монтируется защитная или конструктивная стальная сетка и подшиваются поперечная деревянная обрешетка. Однако, такой способ монтажа возможен только при возможности доступа в подпольное пространство. Что же делать, если высота подпола составляет, например, 40 см? В этом случае монтаж подшивки производится изнутри помещения следующем порядке: |
|
2. После установки балок перекрытий доски обрешетки подшиваются либо саморезами снизу с помощью шуруповерта (рука свободно проходит в промежуток между досками) либо сверху — по два самореза на доску. Также можно использовать крепление на стальных уголках. 3. Стальную сетку и супердиффузионную мембрану укадывают уже сверху по доскам обрешетки. Если вы используете пенопласт или ЭППС, то мембрана не нужна, а подшивка должна быть сплошной — минимально при расстоянии балок 40 см это фанера или OSB-3 толщиной 16 мм. Также, благодаря особым физическим свойствам OSB возможно устройство упрощенных утепленных перекрытий: |
|
|
|
|
|
OSB по результатам современных западных исследований является «умным» паробарьером. это означает, что при влажности 50% паропроницаемость OSB составляет 2 perm (ОСБ не паропроницаемо), а при влажности 85% паропроницаемость ОСБ растет и составляет 12 perm (ОСБ паропроницаемо). 4. Утеплитель укладывается в распор (например плиты Рокувул Лайт Баттс имеют подпружиненные края). Для этого расстояние между балками по внешним сторонам должно быть на 1-2 см уже ширины плиты используемого минераловатного утеплителя. Либо плиты утеплителя нарезаются с напуском на 1 см на сторону. Пенопласт или ЭППС пропенивается по всем щелям. Более толстые плиты укладываются кнаружи. Тонки — кнутри. Швы между плитами должны быть смещены в разных слоях. 5. Поверх утеплителя без зазора укладывается пароизоляция (п/э пленка 150 мк и более, Изоспан Б, фольгированная крафт бумага) с нахлестом листов на 20-30 см и проклейкой стыков скотчем. 6. Поверх пароизоляции без зазора или с зазором (по требованию производителя) укладывается черновой пол + чистовой пол, либо сразу чистовой пол. Толщину плит и досок чернового и чистового пола можно посмотреть вот в этом разделе. |
|
| Что делать если перекрытия у вас уже установлены, уложен пол, а толщина утеплителя недостаточна? | |
|
Вариант А. Я хочу напакостить теще, чтобы пол у нее быстро сгнил. Вариант Б. У меня есть голова на плечах, и я не хочу менять перекрытия еще лет 70. |
|
|
Читать о расчете толщины утепления деревянных полов |
|
| Читать о конструкции теплых полов по деревянным перекрытиям | |
| Читать о конструкции полов по грунту | |
| Теплые полы по деревянным перекрытиям |

 Допустим у вас балки 15 см высотой. Минимально допустимый размер черепных брусков 2,5 см + толщина доски (2 см). Из всей высоты бруса остается лишь 10,5 см на утеплитель. Этого не хватит для эффективного утепления пола минеральной ватой даже если ваш дом находится в Краснодарском крае (минимальная расчетная толщина утеплителя — 14 см). В большинстве случаев, кроме территории Сибири, в России достаточно 20 см толщины минераловатного утеплителя. И как вы будете выкручиваться с черепными брусками? Наращивать балки поверху? Вариант, но это дополнительные расходы и опять таки малые пролеты перекрытия из-за малой высоты несущих балок.
Допустим у вас балки 15 см высотой. Минимально допустимый размер черепных брусков 2,5 см + толщина доски (2 см). Из всей высоты бруса остается лишь 10,5 см на утеплитель. Этого не хватит для эффективного утепления пола минеральной ватой даже если ваш дом находится в Краснодарском крае (минимальная расчетная толщина утеплителя — 14 см). В большинстве случаев, кроме территории Сибири, в России достаточно 20 см толщины минераловатного утеплителя. И как вы будете выкручиваться с черепными брусками? Наращивать балки поверху? Вариант, но это дополнительные расходы и опять таки малые пролеты перекрытия из-за малой высоты несущих балок.


 Мембрана укладывается «корытом» с напуском на стенки балок на 10-15 см и закрепляется скобами степлера. Утеплитель укладывается враспор между балками. Более толстые плиты утеплителя укладываются в нижний слой. Стыки плит в слоях устраиваются в разбежку, чтобы исключить продувание слоя утеплителя.
Мембрана укладывается «корытом» с напуском на стенки балок на 10-15 см и закрепляется скобами степлера. Утеплитель укладывается враспор между балками. Более толстые плиты утеплителя укладываются в нижний слой. Стыки плит в слоях устраиваются в разбежку, чтобы исключить продувание слоя утеплителя.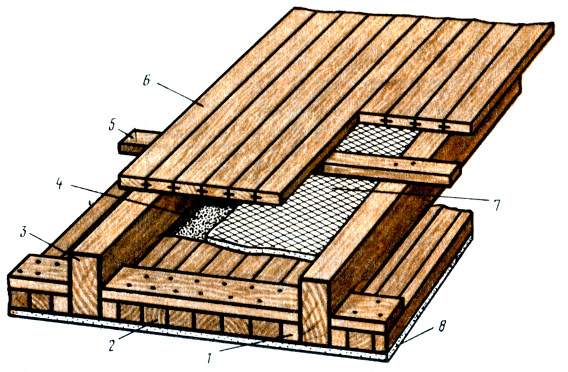
 Предпочтительно использование отражающей пароизоляции: вы сможете сохранить в доме больше тепла за счет отражения его ИК (инфракрасной) — лучистой составляющей.
Предпочтительно использование отражающей пароизоляции: вы сможете сохранить в доме больше тепла за счет отражения его ИК (инфракрасной) — лучистой составляющей.
Брусок — сколько в кубе штук
При подготовке к строительству деревянного здания, дома или постройки будет рассудительнее предварительно узнать общее количество необходимого материала для полного завершения строительства объекта. А когда в процессе используется брусок, то его количество можно узнать при помощи специальных таблиц с расчетами. Или же для этого можно воспользоваться расчетами, проведенными самостоятельными силами.
А когда в процессе используется брусок, то его количество можно узнать при помощи специальных таблиц с расчетами. Или же для этого можно воспользоваться расчетами, проведенными самостоятельными силами.
Математическая модель подсчета количества бруска в кубе
Основной способ подсчета количества бруска в кубе древесины заключается в простой формуле, для этого необходимо площадь прямоугольного поперечника бруска умножить на его длину, а сам поперечник вычисляется умножением высоты на ширину.
Для точного определения количества бруска в кубическом метре древесины, в зависимости от его поперечного сечения и длинны, необходимо произвести расчеты:
Например, для строительства объекта необходимо 100 погонных метров бруска марки 40 на 50 мм, который имеет длину 3 метра. Для этого сначала необходимо узнать площадь его поперечника: 0,04 м * 0,05 м = 0,002 кв. м., она оставляет 0,002 кв. м. Далее необходимо эту площадь умножить на длину бруска: 0,002 * 3 = 0,006 куб. м. – это объем одного бруска с поперечником 40 на 50 мм. Потом делим 1 кубический метр древесины на объем одного бруска: 1 куб. м. / 0,006 куб. м. = 166 штук.
– это объем одного бруска с поперечником 40 на 50 мм. Потом делим 1 кубический метр древесины на объем одного бруска: 1 куб. м. / 0,006 куб. м. = 166 штук.
Вот теперь имеется полученный результат, всего 166 штук бруска в 1-ом (одном) кубическом метре древесины с поперечником 40 на 50 мм и длинной 3 метра. Теперь каждый сможет узнать про брусок – сколько в кубе будет штук брусков. По такой же аналогичной формуле можно производить расчеты количества бруска относительно других видов бруска, имеющего разные показатели и размеры поперечника.
Подсчеты и завершение математической задачи
Из примера, теперь можно подсчитать, что для строительства объекта необходимо будет 100 п. м.* 0,006 куб. м. = 0,6 куб. м. Вот тоже получен результат, для полного завершения строительства объекта, для которого необходимо 100 погонных метров бруска, необходимо купить 0,6 кубических метров древесины. Эта цифра понадобится для пересчета стоимости бруска, купленного поштучно за единицу изделия на стоимость одного куба древесины.
Таблица количества бруска в кубе
| Наименование пиломатериала | Размер сечений бруса (мм) | Сколько бруса (штук) в кубе (в 1 куб. метре) | Объем одного бруса в кубических метрах (м3) |
| Брусок | 25х50х3000 | 266 | 0,0037 |
| Брусок | 30х40х3000 | 277 | 0,0036 |
| Брусок | 30х50х3000 | 222 | 0,0045 |
| Брусок | 40х40х3000 | 208 | 0,0048 |
| Брусок | 50х50х3000 | 133 | 0,0075 |
| Брусок | 50х70х3000 | 95 | 0,01 |
Проект Сосновый бор
Описание проекта дома Сосновый бор
1. Геодезические работы: разбивка осей, нивелировка.
2. Фундамент – буронабивные сваи диаметром 300 мм, L 3000 мм. (32 штуки)
3. Основания – 2 ряда (обвязка) из бруса 200х150мм. Гидроизоляция рубероид.
Гидроизоляция рубероид.
4. Сруб (стены 1-го этажа) — Профилированный брус 190х140мм с полумесяцем, выполнены в полтора этажа
5. Половые лаги (шагом 60 см) — брус 100х150 мм на ребро
6. Сруб собирается (сколачивается) собирается на березовый нагель.
7. Внутренняя и наружная отделка стен, потолка выполнена оцинкованными гвоздями.
8. Сборка углов и длинных стен собирается – В тёплый угол (шип-паз)
9. Между венцами прокладывается льноджутовое полотно
10. Перегородки 1-го этажа — Профилированный брус 90х140мм (прямой). Монтаж в ройку, глубиной 2-4 см.
11.Перегородки 2-го этажа — отсутствует
12. Высота потолка от пола 1-й этаж –2,58 +-3см (21 ряд+ 11 рядов для второго этажа)
13. Высота потолка от пола 2-й этаж –1.26 см продолжения 1-го этажа и 2.30см (общая высота)
14. Крыша –Прямая (по проекту)
15. Высота конька не менее 3.70см
16. В качестве кровли используется гибкая черепица от производителя TEGOLA.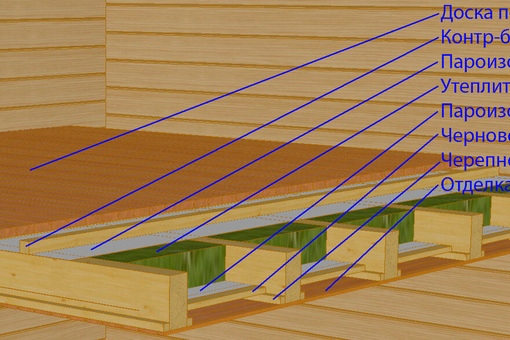 Основа кровли — влагостойкие ОСБ плиты.
Основа кровли — влагостойкие ОСБ плиты.
17.Черновой пол (предназначен для укладки изоляции и утеплителя) – доска обрезная 20 -22мм. Укладывается на черепной брусок 40х50мм-Сорт 1-ый
18. Межэтажные перекрытия (потолочные балки) -брус 150х100 через 60 см на ребро
19. Стропила система из доски 150*40 через 60см.
20. Подрешетник из обрезной доски 100 * 20мм.Сорт 1-ый
21. Стены профилированного бруса не обшиваются.
22 Под подрешетник по стропилам натянута пленка Изеспан D или аналоги (подкровельные плёнки)
23. Фронтоны из проф.бруса 190х140 мм
24. Наружние углы зашиваются хвойной вагонкой
Что такое нагель?
Нагель – это особые штыри (или по-другому шканты), устанавливаемые внутри бруса или бревен при возведении срубов. Установка этих штырей – нагелей необходима для того, чтобы брус или бревно не повело, что неизбежно случалось бы по мере высыхания древесины. Нагеля предотвращают перемещение, смещение бревен относительно друг друга. При правильной установке нагелей исключается возможность кручения бревен или бруса даже если Вы возводите сруб из сырой древесины. Если вы укладываете брус, используя нагеля, ваш дом будет работать только на смятие. Все воздействия, работающие на изгиб, возьмут на себя нагеля.
При правильной установке нагелей исключается возможность кручения бревен или бруса даже если Вы возводите сруб из сырой древесины. Если вы укладываете брус, используя нагеля, ваш дом будет работать только на смятие. Все воздействия, работающие на изгиб, возьмут на себя нагеля.
Чтобы построить качественный брус для круглогодичного проживания, мы использовали нагели цилиндрической формы из березы. Это особенно актуально, если возводится сруб из мягких пород древесины, таких как ель или сосна. Применение березовых нагелей, как правило, обусловлено тем, что сила, возникающая внутри бревен или бруса по мере их высыхания настолько велика, что сруб неизбежно будет скручен и бревно приобретет серповидную форму.
Именно для предотвращения таких процессов мы используем в строительстве березовые нагеля. Так делали наши деды с незапамятных времен, ведь раньше при возведении деревянного дома не использовалось ни одного гвоздя и, следовательно, вся надежда возлагалась только на нагеля. В наши дни считается, что использование нагелей делает деревянную конструкцию более прочной и долговечной. Применение нагелей обеспечивает равномерную усадку сруба.
Применение нагелей обеспечивает равномерную усадку сруба.
Отверстия под деревянный нагель мы сверлим в шахматном порядке. Каждый нагель служит для скрепления между собой двух брусьев. Деревянный нагель вставляется в отверстие, забивается, но не совсем до конца, затем сверху кладется пакля, и сверху накрывается вторым брусом. Только после этого забивается маленьким молоточком.
Что такое лечение Access Bars®?
Как это работает?
Access Bars® помог тысячам людей изменить многие аспекты своего тела и своей жизни, включая сна, здоровье и вес , деньги, секс и отношения, беспокойство, стресс и многое другое.
В худшем случае вы почувствуете, что только что сделали феноменальный массаж. В лучшем случае вся ваша жизнь может с легкостью превратиться во что-то большее.
Некоторые сравнивают мозг с компьютером, а лечение The Bars® — с дефрагментатором диска.Полоски (точки на голове) соответствуют различным областям мозга (компьютерным файлам), которые относятся к аспектам повседневной жизни, таким как радость, творчество, деньги, контроль, создание связей, спокойствие или чувство благодарности и т. Д. собственные ограничивающие мысли, убеждения, эмоции и модели в этих областях (например, ненужные файлы или спам) на основе суждений, полученных от общества, культурной семьи, друзей и личного опыта. Представления о добре и зле, добре и зле мешают принять жизнь такой, какая она есть, и внести изменения, которые наделяют нас силой.Мы можем даже хорошо осознавать, как эти суждения уводят нас от центра, от нашей неотъемлемой связи с целостностью и единством.
Когда практикующий Access Bars® касается прутьев на голове человека, электрический заряд в мозгу, который удерживает эти сфабрикованные суждения, рассеивается (т. Е. Ненужные файлы удаляются!) . Шаблоны и «программы», которые мы взяли, заставляют нас реагировать на жизнь на автопилоте, бессознательно сопротивляясь тому самому опыту, которого мы так жаждем. После очищения мы снова обретаем свободу действовать с ясностью и осознанием, получая то, кем мы являемся на самом деле, без страха и препятствий, и открываемся для совершенно новых перспектив.
Исследования показали, что лечение The Bars® замедляет мозговые волны до тета-состояния, позволяя поведенческим моделям и детским перспективам всплывать и высвобождаться, а также возникает большее состояние присутствия, допущения и открытости . Ограничивающие мысли, убеждения и чувства могут быть сняты, включая страхи по поводу денег, препятствия на пути к творчеству, трудности с получением, остаточное горе или любые другие привычки, которые больше не работают для вас.
The Bars® — это инструмент, который облегчает другой способ функционирования на планете.Триллионы лет человеческие существа функционировали в зависимости от плотности своих мыслей, чувств и эмоций. Это то, с чем люди отождествляли себя, это то, что люди сделали ценным. Чем больше вы запускаете планки, тем больше вы начинаете осознавать, сколько усилий требуется для того, чтобы действовать, исходя из мыслей, чувств и эмоций. Вы также начинаете осознавать возможность функционирования от восприятия, знания, бытия и получения. Для большинства людей запуск своих столбцов — это первый раз, когда они действительно позволяют себе получать без обязательств и внимания.
Если вы чувствуете себя подавленным или не в своем роде, я настоятельно рекомендую попробовать сеанс с Лорой. Это откроет для вас возможности и позволит вам их получить. Настоятельно рекомендуется!Кристал О’Коннелл
Альмонте, Онтарио
Это все равно что активировать компьютерный вирус сознания!
Полоски можно использовать для облегчения изменений во всех сферах вашей жизни. Когда вы желаете действовать с большей осознанностью, вы начинаете открывать двери для всего исцеления. Наука говорит нам, что форма ваших клеток на самом деле становится более эллиптической, когда на них влияют мысли, чувства и эмоции, что является первым шагом к заболеванию.Когда вы запускаете грифы, это разблокирует это воздействие на клетки , позволяя клеткам вернуться к своей более сферической форме, облегчая более легкое пребывание в теле .
В этот момент вы можете подумать (или судить), что это кажется очень «ууууу, Лаура», и я полностью это понимаю. Его эффекты могли быть просто анекдотическими или плацебо — я был там один раз и , пока мои батончики не пробежались в первый раз. Останься со мной … ради настоящей науки.Инструменты Access Consciousness®, в том числе The Bars®, используются психологами, психиатрами, физиотерапевтами, терапевтами для семейных игр, массажистами, иглотерапевтами, мануальными терапевтами, бизнес-тренерами и инструкторами и обычными людьми для создания простых быстрых изменений.
Посмотрите это 11-минутное видео ниже, где нейробиолог д-р Джеффри Л. Фаннин с радостью поделится впечатляющими изменениями, которые он увидел с помощью картирования мозга с термографией, до и после запуска своих столбцов.
Свяжитесь со мной, чтобы узнать больше
Что такое Access Consciousness Bars®?
Хотя нейробиоуправление нейрофидбэком — моя основная специализация, я также практикую Access Bars®, практическую энергетическую модальность. Мягко касаясь определенных точек на голове, этот подход высвобождает электромагнитный заряд мыслей, идей, взглядов и убеждений, которые могли ограничивать вас в соответствующих сферах жизни.
Попросите провести образец сеанса Bars® во время обычного сеанса нейробиоуправления.
На что похож сеанс Bars®?
Все люди разные, поэтому «нормальных» сеансов Bars® не бывает. Чаще всего сообщается о том, что возникает чувство полного расслабления, и многие люди засыпают — даже в очень шумной обстановке. К некоторым людям приходят образы, другие ощущают в своем теле такие ощущения, как жужжание, покалывание, тепло, подергивание, дрожь или даже боль.Многие люди говорят, что энергия, которую они чувствовали, была похожа на удовольствие, которое они получают от возбуждения. Чувство покоя и радости часто присутствует в конце сеанса Bars®.
Это как Рейки?
Нет. Рейки направляет энергии в ваше тело и направляет их на что-то. Слово Рейки состоит из двух японских слов: «Рей», что означает «Мудрость Бога или Высшая Сила», и «Ки», что означает «энергия жизненной силы». Итак, Рейки на самом деле является «духовно управляемой энергией жизненной силы».
Доступ — это разблокирование энергий, которые вы заблокировали в своем теле, чтобы оно высвободилось. Точно так же, как нейробиоуправление ничего не вкладывает в ваш мозг, когда мы запускаем Bars®, мы не вкладываем в вас ничего, чтобы что-то изменить. Мы позволяем ВАМ изменить его, разблокировав место, где вы застряли.
Фасилитатор активирует точки на голове, слегка касаясь их и являясь пространством, в котором могут произойти изменения. В Рейки есть намерение; с Доступом есть надбавка.
Это как EFT?
Нет. EFT — это высвобождение эмоций и травм из тела путем «нажатия» на определенные энергетические точки, и он работает аналогично иглоукалыванию. Это лечение, которое вы можете сделать для себя, чтобы вылечить свое тело. Запуск Bars® и использование инструментов Access выполнят все, что намеревается сделать EFT, и многое другое.
Это похоже на краниосакральную терапию?
Нет, но если вы проведете сеанс Bars®, вы можете получить эффекты, аналогичные краниосакральной терапии.Краниосакральная терапия заключается в мягких манипуляциях с мембраной (фасцией) черепа и вдоль позвоночника, окружающего центральную нервную систему. Это очень точная расслабляющая работа, которая часто вызывает у клиента состояние глубокого расслабления.
Running Bars® вызывает глубокое расслабление для клиента, а также высвобождает энергию мыслей, чувств и эмоций, которые оказали влияние на клетки тела, поэтому вы можете получить больше изменений с Bars®, чем с краниосакральным терапия.
Обратитесь, чтобы забронировать сеанс.Это как рефлексология?
№. Рефлексология основана на том принципе, что, оказывая давление на области стоп, рук и ушей, она снимает напряжение, улучшает кровообращение и способствует естественному функционированию соответствующих участков тела. Bars® не являются массажной техникой и не связаны с точками давления на голове. Однако во время пробежки Bars® может показаться, что вы только что прошли отличный массаж!
Основано ли это на точках акупунктуры, линиях меридианов или чакрах?
№Информация о Bars® была передана в 1990 году и никогда не существовала на планете до этого. Единственное сходство с иглоукалыванием, меридианами и чакрами заключается в том, что Bars® предназначены для работы с энергией.
Если вы знакомы с методами, применимыми к этим энергетическим точкам на теле, вы можете обнаружить, что бег Bars® значительно расширяет возможности любого метода исцеления, в котором они используются. The Bars® здесь, чтобы изменить вашу жизнь. Хотели бы вы это получить?
Больно?
Для активации Bars® давление не требуется.Фасилитатор отрегулирует давление своих рук на вашу голову так, как вам удобно. Были некоторые люди, которым требовалось, чтобы Bars® запускался без прикосновения, и это можно сделать, если у вас чрезмерная чувствительность к прикосновениям. Когда Bars® работают, вы можете испытывать некоторую интенсивность осознания бегущей энергии и можете быть уверены, что она рассеется сама по себе по мере продолжения процесса.
Надо ли снимать одежду?
Нет, фасилитатор будет касаться вашей головы, ладоней и подошв ваших ног.Единственное, что вам, возможно, придется снять, — это обувь и головные уборы, такие как очки и аксессуары для волос, которые могут затруднить доступ к точкам Барс.
Направляет ли Фасилитатор Bars® Дух?
Нет. Ченнелинга не происходит. Фасилитатор ничего не делает, кроме как заставляет бежать энергию каждого бара. Фасилитатор вытягивает энергию через вас, а не от вас, и позволяет этому процессу произвести высвобождение в каждой точке Барс. Фасилитатор будет удерживать позицию до тех пор, пока энергия не потечет плавно, а затем перейдет к следующей точке.
Должен ли я иметь намерение, когда я запускаю свой Bars®?
Нет. Великолепный подарок слитков в том, что они позволяют просто получать. От вас не требуется ничего делать, думать или чувствовать. Вам просто нужно лежать и получать энергию. Это все о вас!
Сколько времени длится сеанс Bars®?
Нет стандартного времени для запуска Баров. Фасилитатор будет следить за энергией и предоставлять только то, что вы счастливы получить. Бары можно сделать всего за 20 минут.Детские батончики бегают быстрее, чем взрослые, так как обычно они меньше сопротивляются приему, чем взрослые.
Могу ли я говорить во время сеанса Bars®?
Да. Вы можете задать вопросы своему фасилитатору или рассказать о том, что для вас происходит. Это не повлияет на сеанс. Вам не нужно сосредотачиваться, медитировать или закрывать глаза.
Могу ли я двигаться во время сеанса Bars®?
Да, конечно. Необходимо, чтобы вы чувствовали себя комфортно, чтобы вы могли легко получить Bars®.Вы можете сделать перерыв в любое время во время сеанса, если вам нужно сходить в ванную, размяться, высморкаться или что-то в этом роде. Вы можете попросить подушку, одеяло или глоток воды. Просите, и вы получите.
Как часто мне следует проводить сеанс Bars®?
Эффект от сеанса Bars® обычно длится около трех недель. Так что все зависит от вас. Многие люди предпочитают получать Bars® один раз в неделю, хотя вы можете делать Bars® каждый день, если хотите. Если вы раздражены или расстроены, это всегда хорошее время, чтобы запустить Bars®.В Калифорнии есть мужчина, который говорит: «Если вы хотите быть счастливым всю оставшуюся жизнь, бегайте каждый день на чьей-нибудь Bars® или на Bars®».
Возбуждены?
Запланируйте сеанс здесьБары были разработаны Гэри Дугласом, основателем Access Consciousness®
Для получения дополнительной информации о Access Consciousness® или Access Bars® посетите веб-сайт Access Consciousness®: https: / /www.accessconsciousness.com/
Краниосакральная терапия, лечебный код и панели доступа
От того, чтобы практикующий работал над исцелением, до обучения методам исцеления, чтобы иметь возможность работать над собой, а затем, наконец, научиться способности присутствовать в своей жизни в каждый момент, без осуждения вас или кого-либо еще, принимать все , ничего не отвергайте и создавайте все, что вы хотите в жизни — больше, чем то, что у вас есть в настоящее время, и больше, чем вы можете вообразить.
Во время сеанса краниосакральной терапии терапевт кладет руки на пациента, что позволяет ему «настроиться на краниосакральный ритм». Практик осторожно воздействует на позвоночник и череп, его черепные швы, диафрагмы и фасции. Таким образом, ограничения нервных проходов уменьшаются, движение спинномозговой жидкости через спинной мозг считается оптимальным, а смещенные кости возвращаются в свое надлежащее положение.
В 2001 году доктор Алекс Лойд открыл, как активировать физическую функцию, встроенную в тело, которая последовательно и предсказуемо устраняет этот единственный источник болезни, чтобы иммунная система тела могла выполнять свою работу по исцелению всего, что в нем не так. Доктор Лойд назвал этот процесс «Кодами исцеления».
Выводы доктора Лойда были подтверждены тысячами людей со всего мира, которые использовали «Коды исцеления» для лечения практически любых физических, эмоциональных или проблем в отношениях, а также для достижения огромных успехов в личном успехе.
Наконец, цель Access Bars — создать мир сознания и единства. Сознание включает в себя все и ничего не судит. Наша цель — довести вас до точки, в которой вы получите от Access осведомленность обо всем без каких-либо суждений. Если у вас нет суждений ни о чем, тогда вы можете смотреть на все так, как оно есть, не за то, что вы хотите, не за то, что должно быть, а просто за то, что оно есть.
Сознание — это способность присутствовать в вашей жизни каждое мгновение, не осуждая вас или кого-либо еще. Это способность получать все, ничего не отвергать и создавать все, что вы желаете в жизни — больше, чем то, что у вас есть сейчас, и больше, чем вы можете вообразить.
Что, если бы вы были готовы заботиться о себе? Что, если бы вы открыли двери для того, чтобы быть тем, кем, по вашему мнению, быть невозможно? Что нужно для вас, чтобы осознать, насколько вы важны для возможностей этого мира?
Частичная кость черепа, найденная в прошлом году, принадлежала человеку с золотым слитком
ЗОЛОТОЙ Слиток — кусок черепа, найденный в прошлом году на берегу реки Скайкомиш, принадлежал человеку с золотым слитком, который пропал без вести с марта 2017 года.
Следователи давно подозревали, что затопленная черепная кость размером с ладонь могла принадлежать 35-летнему Дэниелу Кливленду. На этой неделе его семья узнала, что ответ был подтвержден с помощью ДНК, сообщила медицинская экспертиза округа Снохомиш.
Кливленда в последний раз видели в палаточном лагере к западу от Индекс, чуть выше по течению от песчаной косы, где полтора года спустя остановился кусок черепа. Он считается мертвым.
Кливленд жил с психическим заболеванием.Судебные протоколы показывают, что большую часть последних трех лет своей жизни он провел за решеткой за ненасильственные преступления. Согласно базе данных NamUs, на снимке видно, что он находился в тюрьме за неделю до своего исчезновения.
В то время в публичных бюллетенях отмечалось, что Кливленд «склонен к самоубийству», но точно, как и почему он оказался в реке, до сих пор нет ответа. У него не было машины или мобильного телефона. На нем было темное пальто и джинсы.
Родственники обыскали долину реки весной 2017 года, но не нашли никаких следов Кливленда.
Рыбак заметил кость на берегу реки 10 декабря, на изгибе Неба, в полумиле вниз по течению от Зика Драйв. Власти подтвердили, что это был человек. Других доказательств у них не было. Одежды или других останков не было найдено.
Последние восемь месяцев среди следователей неопознанный мужчина носит прозвище Биг Бенд Джон Доу.
Примерно в прошлое Рождество эксперты сравнили кость с рентгеновским снимком лица, сделанным Кливлендом. Эти двое казались последовательными, но они не были окончательным совпадением.
В мае частная лаборатория в Оклахоме извлекла ДНК. Специалист по генеалогии из Орегона, которая в прошлом работала со следователями округа Снохомиш, Деб Стоун, загрузила генетический профиль на сайт предков GEDMatch, где люди могут искать давно потерянных родственников.
Она начала собирать генеалогическое древо неизвестного мужчины на основе данных о дальних родственниках. Но в середине своего исследования GEDMatch изменила свою политику и по умолчанию ограничила доступ полиции к данным пользователей на сайте, если только пользователи не отметили поле, чтобы сказать, что они согласны с тем, что полиция видит их ДНК.
За ночь количество профилей ДНК, которые она могла использовать для сравнения, упало с 1,2 миллиона до нуля. Стоун обратился к родословным на других сайтах. В итоге она выследила троюродного брата неизвестного мужчины, дважды удаленного по отцовской линии, а затем полу-двоюродного брата по материнской линии.
Имея это подтверждающее свидетельство, судмедэксперт попросил родителей Кливленда сдать анализы ДНК.
Результаты подтвердили, что кость принадлежит черепу Кливленда. Ранее на этой неделе следователи уведомили семью.
В течение последних двух лет друзья и родственники продолжали публиковать сообщения на странице Кливленда в Facebook, прося его вернуться домой, если он жив, и рассказывая ему, как сильно они скучают по нему.
Калеб Хаттон: 425-339-3454; chutton@heraldnet.com. Твиттер: @snocaleb .
Посторбитальная перемычка — обзор
Адаптивное происхождение приматов
Современные приматы отличаются от других млекопитающих, включая большинство плезиадапиформ, многочисленными анатомическими особенностями, в том числе более сходящимися орбитами с заглазничными перемычками и захватом конечностей ногтями, а не когтями на большинстве из них. цифр, а также детали их зубных рядов и анатомии основной черепной коробки.Существует ряд гипотез или сценариев, касающихся конкретных деталей происхождения приматов и поведения, которые были наиболее важными в эволюции этого набора адаптаций приматов (например, Картмилл, 1992; Блох и Бойер, 2002; Солиго и Мартин, 2006; многие статьи в Ravosa and Dagosto, 2007). Эти сценарии часто подчеркивают различные аспекты анатомии приматов и иногда дополняют друг друга.
В гипотезе «прыжка хваткой» Фредерика Салая и Мариана Дагосто (e.g., Szalay and Delson, 1979; Салай и Дагосто, 1980, 1988; Дагосто, 2007; Szalay, 2007), древесные привычки приматов представляют собой примитивную черту евархонтонов, а специфическое поведение, которое отличало верховых приматов от наших предков евархонтонов, включая плезиадапиформ, было более акробатическим типом передвижения с прыжком, который объясняет хватание пальцы и другие особенности конечностей, включая «увеличенные ступни», глубокие коленные суставы и «адаптированную к скорости лодыжку», характерные для самых ранних приматов раннего эоцена.Szalay (2007) отмечает, что ввиду разнообразия морфологии зубов, обнаруженного как у плезиадапиформ, так и у ранних корончатых приматов, невозможно идентифицировать конкретную диетическую привычку, участвующую в происхождении кроновых приматов.
Напротив, Мэтт Картмилл (например, 1970, 1992) уделяет особое внимание визуальной адаптации в происхождении приматов как объяснение черепных особенностей, характерных для приматов, включая заглазничную перемычку, обращенные вперед орбиты и уменьшенные морды. как хватательные руки.Отмечая, что большинство древесных млекопитающих не очень похожи на приматов, поскольку они не обладают ни схождением орбит, ни хватательными пальцами, он утверждает, что мы должны выйти за рамки древесной породы как таковой, чтобы объяснить отличительные черты нашего отряда. Он отмечает, что стереоскопическое зрение особенно характерно для хищников, таких как кошки или совы, которые полагаются на зрение для обнаружения своей добычи. Точно так же он отмечает, что руки большинства древесных млекопитающих оснащены когтями, а не гвоздями, и что прибитые гвоздями хватательные руки приматов, вероятно, являются приспособлением для хватания добычи, а не древесной опорой.Таким образом, в гипотезе «визуального хищничества» Картмилла, предковый примат был особым визуальным хищником, который преследовал и хватал свою добычу в «плотном клубке маленьких веток и виноградных лоз, который характеризует полог и окраину леса» (Cartmill, 1970; Картмилл и др., 2007).
В гипотезе Боба Сассмана (1991, также Sussman et al., 2013) «коэволюция приматов / покрытосеменных» эволюцию приматов лучше всего рассматривать как нечто, происходившее в сочетании с излучением покрытосеменных растений для использования продуктов цветковых растений. (фрукты, цветы и нектар) в небольшой ветке.Хотя он признает, что его теория конкретно не учитывает визуальную адаптацию приматов, он утверждает, что визуальное хищничество — редкое явление среди приматов, которые часто используют звук для определения местонахождения добычи, и вряд ли имело значение для самых ранних приматов. Он также отмечает, что, хотя конвергентные орбиты приматов похожи на орбиты визуальных хищников, таких как кошки и совы, млекопитающие, которые, кажется, показывают наибольшее сходство с приматами по своей зрительной системе, — это фруктовые летучие мыши, группа полностью травоядных.Тем не менее, эволюция покрытосеменных сопровождалась также излучением насекомых, зависящих от этих растений.
Робин Кромптон (1995) подчеркнул роль передвижения, особенно прыжкового поведения у ночных животных, как важного фактора отбора для эволюции стереоскопического зрения приматов, а также посткраниальной адаптации ранних приматов. В его гипотезе о «ночном прыжке» для зрительных и манипулятивных способностей приматов было выбрано движение, а не какой-то конкретный аспект диеты.
Наконец, многие авторы обращались к вопросу о том, вели ли предковые приматы, вероятно, дневной или ночной образ жизни, и большинство данных подтверждают мнение о том, что предковые приматы вели ночной образ жизни (Heesy and Ross, 2001; Ross et al., 2007).
Как подчеркивали многие авторы, ключ к реконструкции истории любого, казалось бы, интегрированного набора эволюционных адаптаций заключается в наличии записи о последовательности, в которой были приобретены особенности. Были ли визуальные адаптации приматов предшествующими или следовавшими за способностями к хватанию и уменьшением бугров коренных зубов? Внешность черт, похожих на приматов, разбросана по другим архонтонам.И Dermoptera, и Scandentia, но не plesiadapiforms, часто имеют орбиты, окруженные костью, как у приматов, но не обращенные вперед. У многих плезиадапиформ есть коренные зубы, напоминающие зубы более поздних приматов, но у них очень специализированные передние зубы. У всех Euarchonta, кажется, есть руки, подходящие для древесного поведения, но ни одна из них не демонстрирует черты задних конечностей и лодыжек, связанные с прыжком, которые характерны для самых ранних кроновых приматов. Как обсуждалось выше, трудно представить себе эволюцию приматов из любой другой группы евархонтонов без радикальных изменений в анатомии и адаптации.
В отсутствие четкой последовательности промежуточных форм в ранней эволюции приматов, несколько авторов обратились к другим группам млекопитающих, особенно сумчатым, чтобы предоставить эволюционные модели ранних адаптаций приматов (Rasmussen and Sussman, 2007). Так, Расмуссен (1990) провел полевое исследование неотропических сумчатых Caluromys derbianus, дидельфид, которые демонстрируют поразительное анатомическое сходство с приматами в том, что имеют относительно большой мозг, почти полную заглазничную перемычку, относительно короткую морду и сходство с приматами. пропорции в его (когтистых) пальцах.Аналогичным образом, при изучении морфологии рук и способностей к хватанию, а также походок сумчатых и просимийских приматов, Caluromys поразительно похож на маленьких просимий, таких как Cheirogaleus и Microcebus (Lemelin, 1999; Lemelin and Schmitt, 2007). ). Поскольку Caluromys более древесный, чем любой из его родственников и кормов для плодов и насекомых среди конечных ветвей, изучение этого сумчатого животного не предлагает никаких доказательств того, что хищничество или плодоядность имеют решающее значение для эволюции черт приматов.Тем не менее, Расмуссен предположил, что визуальные адаптации приматов могли развиться для визуального хищничества у животных, которые уже были приспособлены к добыванию фруктов в конечных ветвях, сценарий, который поддерживает каждое из различных адаптивных объяснений происхождения приматов, но в разное время. К сожалению, до тех пор, пока у нас не будет лучшей летописи окаменелостей из позднего мела и палеоцена и промежуточных форм, предшествующих первому появлению кроновых приматов, детали происхождения приматов останутся скрытыми в тумане времени и филогении.
Функциональные и эволюционные последствия фенестрации черепа у птиц по JSTOR
AbstractСтраусоподобные птицы (Palaeognathae) демонстрируют очень небольшое таксономическое разнообразие, в то время как их родственный таксон (Neognathae) насчитывает примерно 10 000 видов. Основные анатомические различия между двумя таксонами заключаются в черепах. Палеогнатам не хватает элемента в клюве, называемого боковой перемычкой, который присутствует как у предковых теропод, так и у современных неогнатов, и имеет тонкие зоны в костях клюва и крепкие костные элементы на вентральной поверхности их черепа.Здесь мы используем комбинацию моделирования и экспериментов по развитию, чтобы исследовать процессы, которые могли привести к этим различиям. Инженерный анализ методом конечных элементов показывает, что удаление боковых перемычек из неогната увеличивает механическое напряжение в верхнем клюве и вентральных элементах черепа, областях, которые у палеогнатов либо более прочные, либо более гибкие. Хирургическое удаление боковой перемычки у новорожденных детенышей привело к аналогичным изменениям. Эти результаты указывают на то, что латеральный стержень является несущим, и предполагают, что эта функция была передана другим костным элементам, когда она была потеряна у палеогнатий.Возможно, что потеря несущей боковой перемычки могла ограничить диверсификацию морфологии черепа палеогнатий и, таким образом, ограничить таксономическое разнообразие внутри группы.
Журнал InformationEvolution, публикуемый для Общества изучения эволюции, является ведущим изданием, посвященным изучению органической эволюции и интеграции различных областей науки, связанных с эволюцией. В журнале представлены важные и оригинальные результаты, которые расширяют наше понимание эволюционных явлений и процессов.
Права и использование Этот предмет является частью коллекции JSTOR.
Условия использования см. В наших Положениях и условиях
© 2017 Автор (ы). Эволюция © 2017 Общество изучения эволюции
Запросить разрешения
эволюция черепной коробки и производительности у чешуекрылых: Последствия редукции черепа и кости для пищевого поведения | Интегративная и сравнительная биология
Аннотация
Эволюция черепной коробки лепидозавров характеризуется общей тенденцией к утрате черепных элементов.Развитие относительно более легких черепов у чешуекрылых, по-видимому, тесно связано с уменьшением относительной массы аддуктора челюсти, что подразумевает функциональные последствия для силы укуса и поведения при кормлении. Интересно, что у чешуйчатых животных заглазничная перемычка была уменьшена или утрачена по крайней мере дважды независимо друг от друга, а таксоны, для которых характерна утрата этих черепных элементов (например, гекконы и вараниды), как правило, имеют подвижный череп. У Gekkotans потеря заглазничной перемычки сопровождалась уменьшением надвисочной перемычки, что приводило к выраженному краниальному кинезису.Наши данные показывают, что кинетический череп имеет функциональные последствия и приводит к снижению силы укуса. Более низкая сила укуса, в свою очередь, может быть причиной снижения эффективности кормления, что отражается в более длительной продолжительности внутриротовых транспортных циклов. Однако геккотанцы, похоже, используют свою внутричерепную подвижность таким образом, что увеличивает скорость движения челюсти при открытии и закрытии, что может позволить им захватывать более неуловимую добычу. Морфологические изменения, наблюдаемые в эволюции черепной системы у чешуекрылых, по-видимому, тесно связаны с функциональными и конструктивными требованиями к черепу, что делает эволюцию плоского черепа модельной системой для исследования последствий морфологических изменений в сложной интегрированной системе производительности, поведения и экология.
Введение
Черепная система позвоночных — яркий пример сложной интегрированной системы. Черепная система состоит из множества высоко интегрированных структурных единиц (например, гиолингвальной системы, челюстей, мозга и органов чувств), каждая из которых связана с различными функциональными и структурными требованиями, предъявляемыми к системе в целом, и формируется ими. Хотя различные компоненты черепной системы имеют решающее значение во многих экологических и поведенческих контекстах, использование челюстей и языка для кормления (обзор Schwenk 2000), питья (обзор Bels et al.1994), а боевое поведение (Lailvaux et al. 2004; Huyghe et al. 2005; Lappin and Husak 2005; Lappin et al. 2006), по-видимому, имеет первостепенное значение для фитнеса. Однако другой важной функцией черепной системы является размещение и защита мозга и основных сенсорных систем. Таким образом, конкурирующие потребности в пространстве (например, Barel 1983) и функциональности (Herrel et al. 2001b), вероятно, будут определять эволюцию системы в целом. Поскольку не все функции могут быть оптимизированы одновременно (например, большие глаза у ночных хищников занимают пространство, которое может уменьшить пространство, доступное для мышц челюсти), черепная система является продуктом конфликтующих избирательных давлений, приводящих к функциональным компромиссам.Таким образом, черепная система позвоночных на первый взгляд может показаться сильно ограниченной в процессе эволюции.
Некоторые группы позвоночных, такие как лепидозавры ( Sphenodon , ящерицы и змеи), демонстрируют поразительное разнообразие строения черепа, что делает череп лепидозавра идеальным тестовым примером для изучения роли функциональных компромиссов и конструктивных ограничений ( sensu Барел 1983) в сложных интегрированных системах. Например, кормление крупной, твердой или жесткой добычей требует более высоких характеристик укуса.Увеличение силы укуса может происходить только за счет (1) увеличения мышечной массы аддуктора челюсти, (2) изменений в архитектуре или физиологии мышц (например, перистых мышц с короткими волокнами; экспрессия различных типов волокон) или (3) изменений в мышечной структуре. механика рычажной системы (Herrel et al. 2002a, 2002b). Если животные не укорачивают внешнюю часть челюстной системы и кусают ближе к точке опоры, увеличение мышечной силы и силы укуса приведет к увеличению нагрузки на черепные элементы, которые должны выдерживать эти большие силы.Таким образом, ожидается, что увеличение силы укуса и мышечной массы будет коррелировать с повышенной прочностью черепа. Тем не менее, механические системы, оптимизированные для создания силы, нельзя одновременно оптимизировать по скорости из-за ограничений на механику рычага и физиологию мышц. Из-за этого ожидается, что виды, охотящиеся на быструю, неуловимую добычу, будут иметь длинные морды, длинные параллельные волокнистые мышцы и длинные внутренние рычаги, открывающие челюсти.
Как упоминалось ранее, не только функциональные требования, но и конструктивные ограничения могут стимулировать эволюцию черепа, как было ранее продемонстрировано для чихлид (Barel 1983).Например, пространство, занимаемое аддукторами челюсти, нельзя использовать для органов чувств или центральной нервной системы, если общий объем черепа должен оставаться неизменным. Кроме того, у некоторых групп ящериц может присутствовать сильное преобладание определенных сенсорных систем, что может оказывать определенное избирательное давление на черепную систему. Например, животные, в значительной степени полагающиеся на химиочувствительность, имеют удлиненные раздвоенные языки с относительно небольшой площадью поверхности языка и длинными мордами. Таким образом, язык больше не может использоваться для транспортировки добычи, и необходимо разработать альтернативные способы транспортировки.У змей это привело к хорошо известной высококинетической системе челюстей, которая используется для транспортировки добычи через ротовую полость (Gans 1961). Точно так же у ночных животных, таких как гекконы, преобладают зрительные и обонятельные чувства, а глаза намного больше, чем у других ящериц. Увеличение размера глазного яблока (Werner and Seifan 2006) вероятно связано с потерей заглазничной и надвисочной перемычки у этих животных (Herrel et al. 1999c, 2000). Очевидно, что потеря этих черепных элементов будет иметь функциональные последствия для стабильности и прочности черепа и повлияет на использование системы во время кормления.
В других группах уменьшение черепных элементов, по-видимому, связано с привычками проживания в щелях (Arnold 1998). Была выдвинута гипотеза, что плоские головы и подвижные черепа кордилид (Cooper et al. 1999), ксенозавров (Ballinger et al. 1995; Lemos-Espinal et al. 1996; Herrel et al. 2001c) и некоторых ласертид (Arnold 1998) ) позволяют животным влезать в расщелины в качестве тактики борьбы с хищниками. Таким образом, разные факторы отбора могут привести к одинаковым функциональным требованиям и могут иметь одинаковые функциональные последствия в разных группах.Тем не менее, поскольку предок Bauplan может отличаться для этих групп, аналогичные функциональные требования потенциально могут также привести к альтернативным решениям. Функциональная конвергенция не обязательно должна быть связана со сходными морфологическими изменениями в основных компонентах системы в разных группах.
Здесь мы применяем комплексный подход к эволюции черепной структуры у ящериц, исследуя морфологические основы, а также функциональные и поведенческие последствия изменений морфологии.В качестве первоначального подхода мы исследуем, действительно ли общая прочность черепа связана с размером челюсти и приводящих мышц, как предсказывалось ранее. Если это так, то изменение прочности черепа и потеря черепных костей могут иметь серьезные функциональные последствия для силы укуса, производительности при кормлении и, в конечном итоге, для экологии животных. Мы используем поразительное сокращение черепных костей у гекконов и других групп ящериц в качестве тестового примера для изучения функциональных последствий изменений морфологии для функций, поведения и экологии.
Материалы и методы
Животные
Образцы для препарирования (Таблица 1) были животными, которые умерли во время транспортировки для коммерческой торговли домашними животными. Они хранятся в Лаборатории функциональной морфологии Университета Антверпена. Образцы, использованные для анализа силы укуса, были собраны в полевых условиях, протестированы и выпущены в течение 24 часов после захвата. Все образцы, снятые для количественной оценки пищевого поведения, были получены в рамках коммерческой торговли домашними животными.
Таблица 1Мышечные массы черепа и челюсти
90339033
9033 903 30 7,11
9033 cuvieri flavimaculata| Семья . | Род . | Виды . | Длина черепа (мм) . | Масса черепа (г) . | Масса аддуктора (мг) . | MPPt (мг) . | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Agamidae | Agama | stellio | 24.00 | 0,53 | 1250,00 | 10,00 | |||||||
| Agamidae | Pogona | vitticeps | 35,21 | 1,30 331 | 3 3565323 | 3 3565303 9033io acanthinurus | 32,44 | 2,06 | 838,28 | 8,45 | |||
| Amphisbaenia | Blanus | cinereus | 8.08 | 0,02 | 16,59 | ||||||||
| Anguidae | Gherronotus | infernalis | 28,77 | 28,77 | 0,39 | 37530 | 0,39 | 37530 9034 | 36,50 | 2,00 | 1864,64 | ||
| Cordylidae | Cordylus | тропидостернон | 23.55 | 0,26 | 155,38 | 2,39 | |||||||
| Corytophanidae | Василиск | Basiliscus | 28,31 | 0,36 | 438,37 | 4,21 | |||||||
| Crotaphytidea | Crotaphytus | collaris | 26,63 | 0,89 | 444,29 | 6,14 | |||||||
| Eublepharidea | Eublepharis | macularius | 29.85 | 0,74 | 695,91 | 23,97 | |||||||
| Gekkonidae | Gekko | gecko | 36,71 | 1.01 | 1.01 | 21 madagascariensis | 25,68 | 0,39 | 354,60 | 12,90 | |||
| Gherrosauridae | Gherrosaurus | major | 36.00 | 1,64 | 379,30 | 3,64 | |||||||
| Helodermatidae | Heloderma | suspectum | 45,00 | 7,11 | 1 | игуана | 37,23 | 0,83 | 349,41 | 5,15 | |||
| Lacertidae | Gallotia | galloti 30.47 | 0,76 | 724.03 | 4,48 | ||||||||
| Lacertidae | Podarcis | atrata | 15,55 | 0,09 | 29,46 | 0,60 | 332,89 | 4,50 | |||||
| Phrynosomatidae | Phrynosoma | douglassi | 42 | 0,29 | 216,31 | 1,42 | |||||||
| Phrynosomatidae | Phrynosoma | platyrhinos | 15,86 | 0,24 | 70,18 | 1,06 | |||||||
| анолисовые | Anolis | garmani | 34,32 | 0,62 | 415,98 | 7,27 | |||||||
| Scincidae | Corucia | zebrata | 51.10 | 3,57 | 4650,00 | 30,00 | |||||||
| Scincidae | Novoeumeces | schneideri | 28,14 | 0,56 | 1 9033 9033 9033 | 0,56 | 1 9034 fernandi | 25,22 | 0,33 | 290,69 | 7,83 | ||
| Scincidae | Tiliqua | scincoides | 65.80 | 7,38 | 10170,00 | 90,00 | |||||||
| Змеи | Nerodia | fasiata | 22.09 | 0,433 | 264.60 331 9033 | 263 ameiva | 40,46 | 1,59 | 1511,44 | 3,59 | |||
| Tropiduridae | Leiocephalus | carinatus | 21.43 | 0,25 | 134,88 | 2,87 | |||||||
| Varanidae | Varanus | niloticus | 38,94 | 0,44 | 3 12301 | 0,44 | 3 12301 | 0,44 | 3 12301 | 19,76 | 0,19 | 68,02 | 0,73 |
| Семья . | Род . | Виды . | Длина черепа (мм) . | Масса черепа (г) . | Масса аддуктора (мг) . | MPPt (мг) . | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Agamidae | Agama | stellio | 24,00 | 0,53 | 1250,00 | 10,00 | 1,30 | 3565,25 | 17,13 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Leiolepididae | Uromastix | acanthinurus | 32,44 | 2,06 | 838,28 | 8,45 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| двуходки | бланусы | cinereus | 8,08 | 0,02 | 16,59 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Anguidae | Gherronotus | infernalis | 28.77 | 0,39 | 375,31 | 5,44 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Chameleonidae | Chameleo | calyptratus | 36,50 | 2,00 | 1864,64 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cordylidae | Cordylus | tropidosternon | 23,55 | 0,26 | 155,38 | 2,39 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Corytophanidae | Basiliscus | basiliscus | 28.31 | 0,36 | 438,37 | 4,21 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Crotaphytidea | Crotaphytus | collaris | 26,63 | 0,83 444 | 26,63 | 0,83 444 | macularius | 29,85 | 0,74 | 695,91 | 23,97 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gekkonidae | Gekko | gecko | 36.71 | 1,01 | 2142,90 | 66,90 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gekkonidae | фельзумы | madagascariensis | 25,68 | 0,39 | 354,60 | 12,90 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gherrosauridae | Gherrosaurus | major | 36,00 | 1,64 | 379,30 | 3,64 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Helodermatidae | Heloderma | suspectum | 45.00 | 7,11 a | 3930,00 | 34,00 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Iguanidae | Iguana | Iguana | 37,23 | 0,83 | 37,23 | 0,83 | 37,23 | 0,83 | галлот | 30,47 | 0,76 | 724,03 | 4,48 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lacertidae | Podarcis | atrata 15331 | 0,09 | 104,60 | 2,00 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Opluridae | Oplurus | cuvieri | 29,46 | 0.60 | douglassi | 17,42 | 0,29 | 216,31 | 1,42 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phrynosomatidae | Phrynosoma | platyrhinos | 0,24 | 70,18 | 1,06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Polychrotidae | Anolis | garmani | 34,32 | 0,62 | 1 9033 9033 9033 9033 Corida | 0,62 | 1 9033 9034 zebrata | 51,10 | 3,57 | 4650,00 | 30,00 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Scincidae | Novoeumeces | schneideri | 28.14 | 0,56 | 259,54 | 3,51 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Scincidae | Riopa | Fernandi | 25,22 | 3 | 2 | 65,80 | 7,38 | 10170,00 | 90,00 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Serpentes | Nerodia | fasciata | 22.09 | 0,42 | 264,60 | 67,03 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Teiidae | Ameiva | ameiva | 40,46 | 1,59 | 1 | 21,43 | 0,25 | 134,88 | 2,87 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Varanidae | Varanus | niloticus | 38.94 | 0,44 | 262,01 | 12,89 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Xantusiidae | Лепидофима | flavimaculata | 19,76 | 9301 | 19,76 | 3 0,19 | 19,76 | 3 0,19 | череп и челюсть Agamidae 30 90335586 9033 0,33 scincoides 903 carinatus Agamidae 30 9033 9033 9033 9033 9033 9033 9033 9033 9033 fasiata
МорфологияВсе образцы, использованные для анализа мышечной массы, хранили в 10% -ном водном растворе формальдегида в течение 24–48 часов, в зависимости от размера образца. После фиксации образцы промывали водой и переводили в 70% водный раствор этанола. Все образцы хранились в этаноле не менее двух месяцев перед вскрытием, что обеспечивало аналогичную степень обезвоживания ткани.Все черепные мышцы удаляли из образцов и хранили в 70% этаноле до взвешивания. Мышцы промокали насухо и взвешивали на электронных весах Mettler MT5 (точность: ± 0,01 мг). Поскольку спирт непрерывно испарялся из мышц даже после того, как он был высушен, через 10 с регистрировали массу всех мышц. Черепа и нижние челюсти были вручную очищены от оставшихся мягких тканей после удаления черепных мышц и взвешены отдельно на электронных весах. Данные о мышечной массе некоторых видов взяты из ранее опубликованных работ (Herrel et al.1996b, 1998, 1999a, 2000), а при необходимости собирались дополнительные данные. Усилие прикусаСилы укуса были получены для ящериц из одной пустынной общины в Южной Австралии. Ящериц ловили руками и заставляли укусить изготовленный на заказ регистратор силы укуса, созданный из преобразователя силы Kistler 9203, присоединенного к портативному усилителю заряда Kistler 5058A (описание см. В Herrel et al. 1999a, 2001a, 2001c). аппарат). После сбора данных о силе укуса ящерицы были измерены (длина морды и выступа; размеры головы) и возвращены к точному месту отлова в течение 24 часов. Кинематика и поведение кормленияЯщерицы, использованные для испытаний пищевого поведения (список видов см. В Таблице 2), были получены через коммерческую торговлю домашними животными и содержались в лаборатории функциональной морфологии Университета Антверпена. Им давали воду и пищу ad libitum и поддерживали 12-часовой цикл свет / темнота. Для всех ящериц было предусмотрено место для купания при более высокой температуре (45–50 ° C), что позволяло им достичь желаемой температуры.Перед съемками ящериц поместили в акриловую клетку с узким выступающим коридором и преподнесли ряд продуктов. Для всех насекомоядных и всеядных видов были собраны данные о сверчках, кузнечиках и мучных червях в качестве добычи. Для травоядных и всеядных ящериц в качестве дополнительных продуктов питания были включены бананы, помидоры и эндивий (Herrel et al. 1996a, 1999b, 1999c; Herrel and De Vree, 1999). Таблица 2Кинематика и пищевое поведение различных ящериц и змей 1848551 9033 9033 P0331 9033 PAL9055 903 903
Кинематическое и пищевое поведение различных ящериц и змей 9033 9033 P0331 9033 9033 P0331 9033309033 Sceloporus
АнализыВсе данные были преобразованы в журнал 10 перед анализом. Регрессия была проведена для изучения взаимосвязи между размером черепа и мышечной массой или силой укуса. В качестве независимой переменной был выбран размер головы, а не длина рыла-отверстия (традиционная мера размера тела у лепидозавров) из-за больших различий в форме тела среди таксонов (например, у змей и безногих ящериц чрезвычайно удлиненные тела) и из-за того, что отношение к пищевому поведению.Поскольку виды или группы видов не являются независимыми точками данных, исторический характер данных принимался во внимание при изучении взаимосвязей между переменными путем вычисления независимых контрастов (Felsenstein 1985). При анализе использовались две радикально расходящиеся гипотезы о взаимоотношениях между основными группами ящериц. Первое — это консенсусное дерево, в значительной степени основанное на морфологических данных, которое ставит игуанов (акродонтов + Iguanidae s.l.) базальными по отношению ко всем другим ящерицам и ящериц-ксантусид в качестве сестринского таксона ласертиформ (Estes et al.1988) (рис. 1А). Отнесение змей к таксону-сестре варанид основано на работе Ли (1998). Второе дерево — это снова дерево консенсуса, но основанное на молекулярных данных. Этот анализ помещает геккосов базальнее всех других ящериц и помещает ксантусид и кордилид в качестве сестринских таксонов сцинцидам (Vidal and Hedges 2004; Townsend et al. 2004; Fry et al. 2005; Fig. 1B). Учитывая убедительные доказательства сестринских отношений между лацертидами и амфисбенидами (Harris et al., 2001; Vidal, Hedges, 2004; Townsend et al.2004; Фрай и др. 2005) во всех анализах они считались сестринскими таксонами. Внутригрупповые отношения были установлены согласно Metzger and Herrel (2005). Поскольку мы не смогли установить время дивергенции для всех видов в анализе, длина всех ветвей была принята равной единице (Мартинс и Гарланд, 1991; Диас-Уриарте и Гарланд, 1998). Мы использовали пакет PDAP (Гарланд и др., 1999) для нашего анализа. Мы проверили диагностические графики и статистику в программе PDTREE, чтобы убедиться, что наши постоянные длины ветвей соответствуют всем признакам (Garland et al.1999). Рис. 1 Филогенетическое древо, отображающее отношения между основными кладами лепидозавров на основе морфологических данных (см. «Методы»). Справа от морфологического дерева консенсуса показаны изображения черепов репрезентативных видов каждой группы. Serpentes, Nerodia fasciata ; Varanidae, Varanus bengalensis ; Anguidae, Gerrhonotus infernalis ; Teiidae, Ameiva ameiva ; Lacertidae, Gallotia galloti ; Amphisbaenidae, Blanus cinereus ; Xantusiidae, Lepidophyma flavimaculata ; Scincidae, Corucia zebrata ; Cordylidae, Cordylus tropidosternon ; Gekkonidae, Eublepharis macularius ; Iguanidae sensu lato , Anolis garmani ; Acrodonta, Plocederma stellio ; Sphenodontidae, Sphenodon punctatus . Рис. 1 Филогенетическое древо, отображающее отношения между основными кладами лепидозавров на основе морфологических данных (см. «Методы»). Справа от морфологического дерева консенсуса показаны изображения черепов репрезентативных видов каждой группы. Serpentes, Nerodia fasciata ; Varanidae, Varanus bengalensis ; Anguidae, Gerrhonotus infernalis ; Teiidae, Ameiva ameiva ; Lacertidae, Gallotia galloti ; Amphisbaenidae, Blanus cinereus ; Xantusiidae, Lepidophyma flavimaculata ; Scincidae, Corucia zebrata ; Cordylidae, Cordylus tropidosternon ; Gekkonidae, Eublepharis macularius ; Iguanidae sensu lato , Anolis garmani ; Acrodonta, Plocederma stellio ; Sphenodontidae, Sphenodon punctatus . Чтобы проверить, имеют ли виды с явно кинетическим черепом (как показывают наши кинорадиографические данные, т. Е. Гекконы, Cordylus и змеи) более крупные мышцы constrictor dorsalis , чем у других ящериц (Таблица 1), был проведен анализ моделирования с использованием программы PDSIMUL и PDANOVA (Гарланд и др., 1993). В программе PDSIMUL мы использовали броуновское движение в качестве нашей модели эволюционных изменений и выполнили 1000 неограниченных симуляций, чтобы создать эмпирическое нулевое распределение, с которым можно было бы сравнить значение F из исходных данных.В программе PDANOVA в качестве фактора был введен кинетизм черепа, масса м. protractor pterygoidei (MPPt) использовался как независимая переменная, а длина черепа — как ковариант. Мы считали различия между категориями значимыми, если исходное значение F было выше, чем значение F 95 , полученное из эмпирического распределения на основе моделирования. Опять же, был проведен анализ двух наборов филогенетических отношений между таксонами (см. Выше). Для проверки различий между группами в кинематике и поведении при кормлении (таблица 2) использовались традиционные ANCOVA. Филогенетически обоснованный анализ этих данных не проводился, поскольку сравнивались только несколько групп с радикально расходящейся морфологией черепа (например, гекконы по сравнению со всеми другими ящерицами). В это сравнение не были включены никакие травоядные виды. Чтобы избежать смещения результатов из-за влияния типа добычи, использовались только данные для ящериц, питающихся аналогичной добычей (сверчки, кузнечики и мучные черви).Обратите внимание, однако, что змей кормили золотых рыбок во время испытаний кормления. Результаты и обсуждениеМорфология черепа ящериц удивительно разнообразна и, по-видимому, характеризуется повторяющейся независимой потерей черепных элементов в некоторых линиях (рис. 1 и 2; таблица 1). По сравнению с состоянием базального диапсида, представленным Sphenodon , плоскоклеточные характеризуются отсутствием квадратично-скуловой кости и потерей нижней височной перемычки (Rieppel and Gronowski 1981).Уменьшение нижней височной перемычки и открытие нижнего височного отверстия, вероятно, произошло до разделения чешуек и ринхоцефалов (Muller 2003) и, по-видимому, связано с расширением внешней приводящей мышцы к латеральной стороне нижней челюсти ( Риппель и Гроновски 1981). Рис. 2 Филогенетическое дерево на основе молекулярных данных (см. «Методы»). Обратите внимание на основные различия в положении геккотанов и игуанов на двух деревьях.Справа изображена эволюция морфологического разнообразия черепной формы. Рис. 2 Филогенетическое древо на основе молекулярных данных (см. «Методы»). Обратите внимание на основные различия в положении геккотанов и игуанов на двух деревьях. Справа изображена эволюция морфологического разнообразия черепной формы. Среди чешуйчатых верхняя височная перемычка утрачена только у гекконов, змей и амфисбенов. Несмотря на предположительное базальное положение гекконов среди чешуйчатых (на основании молекулярной филогении) (рис.2), это, по-видимому, уникальная и производная черта для группы. Радикально расходящаяся морфология черепа у змей и амфисбенов также является уникальной и происходит от обеих групп и может быть связана с окаменелостями в существующих или предковых формах. У других чешуек, таких как большинство варанидов, некоторых ангидов (например, Gherronotus ) и некоторых тейидов (например, Aspidoscelis ), верхняя височная перемычка стала намного тоньше и обеспечивает прикрепление только к апоневрозу поверхностной приводящей мышцы.У других плоскоклеточных клеток верхняя височная перекладина также функционирует как место прикрепления волокон поверхностной части внешнего аддуктора. Среди непрыгающих форм и вараниды, и гекконы дополнительно характеризуются уменьшением заглазничной перемычки. Таким образом, несмотря на гипотезу о том, что черепная система должна быть сильно ограничена в эволюции, кажется, что присутствует множество вариаций. Это поднимает вопрос о том, какие функциональные ограничения могут влиять на черепную конструкцию и различаются ли они в разных кладах ящериц.Одна из гипотез, выдвинутых во введении, заключается в том, что черепная система, вероятно, ограничена своей функцией во время кормления и генерацией силы укуса в целом (например, во время кормления, борьбы и защитного поведения). С механической точки зрения можно ожидать, что череп должен быть сконструирован таким образом, чтобы минимально выдерживать силы реакции на пищу и прямые мышечные силы, прилагаемые в местах происхождения и прикрепления. Таким образом, ожидается корреляция между прочностью черепа и рабочим режимом нагружения при различных формах поведения, что может наложить ограничение «минимальный дизайн» на конструкцию черепа. Чтобы разобраться в этом вопросе, были сопоставлены мышечные массы и размер черепа у большинства линий ящериц. Если минимальные критерии дизайна влияют на эволюцию черепа у ящериц, тогда должна присутствовать связь между размером черепа и силами. Поскольку практически невозможно оценить режим нагрузки in vivo и для широкого круга представителей разных размеров, мы использовали массу закрывающей мускулатуры челюсти как показатель максимальной силы, которая может быть приложена к черепу.Наш анализ показывает, что как масса черепа, так и масса аддуктора челюсти сильно коррелируют с длиной черепа (регрессия стандартизованных независимых контрастов с использованием морфологического дерева: R 2 = 0,83; P <0,001; с использованием молекулярного дерева: R 2 = 0,84; P <0,01). Таким образом, не удивительно, что эволюция более крупной головы связана с эволюционным увеличением массы приводящей мышцы челюсти. Однако более интересным является то, что прочность черепа (т.е., остатки независимых контрастов массы черепа над независимыми контрастами длины черепа) значимо связаны с остаточными контрастами массы челюстно-приводящей мышцы (морфологическое дерево: r = 0,52, P = 0,004, рис. 3A; молекулярное дерево: r = 0,53, P = 0,003, рис. 3B). Таким образом, виды с более массивным и крепким черепом имеют относительно большие приводящие челюсти. Таким образом, наша гипотеза о минимальном ограничении конструкции подтверждается данными и предполагает, что укус действительно формирует эволюцию конструкции черепа и всей черепной системы. Рис. 3 Взаимосвязь между прочностью черепа и размером приводящей мышцы челюсти у ящериц. Ящерицы с относительно более тяжелым черепом (относительно длины черепа) имеют относительно большие приводящие мышцы челюсти. ( A) Анализ на основе морфологического дерева, изображенного на рис. 1. ( B) Анализ на основе молекулярного дерева, изображенного на рис. 2. Рис. 3 Взаимосвязь между прочностью черепа и размером приводящая челюсть у ящериц.Ящерицы с относительно более тяжелым черепом (относительно длины черепа) имеют относительно большие приводящие мышцы челюсти. ( A) Анализ на основе морфологического дерева, изображенного на рис. 1. ( B) Анализ на основе молекулярного дерева, изображенного на рис. 2. Предыдущий анализ черепной массы у ящериц (Metzger and Herrel 2005) предполагал что у травоядных ящериц относительно легкий череп. С другой стороны, анализ силы укуса у травоядных и всеядных животных показывает, что они кусают сильнее, чем другие ящерицы (Herrel et al.2004 г.). Хотя это может означать, что эти виды обошли требования к дизайну черепа, налагаемые режимом нагрузки, это кажется маловероятным, учитывая общность наших результатов, основанных на наборе данных, который включал несколько травоядных и всеядных видов (например, Iguana iguana , Pogona vitticeps , Uromastix acanthinurus , Gallotia galloti , Podarcis atrata и Corucia zebrata ). Интересно, что анализ формы черепа также предполагает, что виды, которые включают в свой рацион растительные вещества, имеют относительно короткие морды (Metzger and Herrel 2005).За счет уменьшения длины рыла длина внешнего рычага челюсти минимизируется по сравнению с закрывающим рычагом, что увеличивает механическое преимущество системы без увеличения мышечных сил. Тем не менее, у некоторых специализированных травоядных также были продемонстрированы мышечные адаптации, включая увеличение мышечной массы (Herrel et al. 1998). Тщательное изучение черепов некоторых из этих видов (например, Corucia zebrata и Gallotia galloti ) позволяет предположить, что черепа укреплены дермальными остеодермами, которые не учитываются при оценке черепной массы.У других видов больший момент закрытия челюсти вокруг челюстного сустава достигается без увеличения мышечной силы, прежде всего за счет изменения ориентации мускулов челюсти (Herrel et al. 1998). Таким образом, травоядные и всеядные ящерицы, по-видимому, также соответствуют минимальным критериям проектирования и имеют черепа, построенные так, чтобы выдерживать режим нагрузки, налагаемый мышечными силами приводящих челюстей. Очевидно, что мышечные силы — не единственное ограничение, ограничивающее конструкцию черепной системы, и у некоторых ящериц (например.g., geckos) дизайн черепа был связан с наличием хорошо развитой сенсорной системы. Ранее предполагалось, что ночной образ жизни общего предка гекконов привел к увеличению размера глаз, как это наблюдается у современных гекконов (Werner and Seifan 2006). Из-за ограничений физического пространства увеличение размера глаза привело к потере посторбитальных и надвисочных черепных элементов, что, вероятно, привело к выраженной внутричерепной подвижности у этих животных (Herrel et al.2000). Таким образом, пространственные ограничения могут потенциально влиять на дизайн черепа у ящериц. Однако должны быть соблюдены минимальные критерии проектирования, и пространственные ограничения, действующие на черепа гекконов, могут иметь серьезные последствия для режима максимально допустимой нагрузки и, следовательно, также способности укусить этих животных. Чтобы изучить этот вопрос, мы проанализировали данные о силе укуса ящериц из одного сообщества пустынных ящериц в Австралии. Наши данные показывают, что гекконы действительно имеют значительно меньшую силу укуса для их размера головы (длины головы), чем у агамидных или сцинцидных ящериц из того же сообщества (ANCOVA: F 2,129 = 39.9; P <0,001; Рис.4). Учитывая, что этот анализ основан на уровне сообщества, мы можем оценить влияние дизайна черепа на разделение ресурсов в сообществе. Из-за их более низкой силы укуса гекконы потенциально могут быть ограничены в их рационе, поскольку было продемонстрировано, что сила укуса связана с эффективностью кормления и шириной рациона по функциональным осям (то есть размером жертвы, твердостью и уклончивостью) (Verwaijen et al. 2002; Херрел и др. 2006). Тем не менее, из-за своих ночных привычек гекконы, вероятно, не имеют или почти не имеют конкуренции со стороны дневных ящериц и могут специализироваться на более мягкой доступной добыче. Рис. 4 График, отображающий взаимосвязь между длиной головы и силой укуса ( R 2 = 0,92; P <0,001; наклон = 2,35; точка пересечения = - 1,39) для ящериц одного сообщества пустынных ящериц из Австралии. Обратите внимание на то, что гекконы кусают значительно меньше для своего размера головы, чем агамиды или сцинциды, согласно анализу ковариации. Рис. 4 График, отображающий взаимосвязь между длиной головы и силой укуса ( R 2 = 0,92; P <0,001; наклон = 2,35; точка пересечения = - 1,39) для ящерицы единого сообщества пустынных ящериц из Австралии.Обратите внимание на то, что гекконы кусают значительно меньше для своего размера головы, чем агамиды или сцинциды, согласно анализу ковариации. Тем не менее, гекконы — не единственные ящерицы, у которых, как считается, есть кинетические черепа, и поэтому они могут столкнуться с теми же ограничениями силы укуса (см. Обзор в Metzger 2002). Однако удивительно, что исследование пищевого поведения ящериц с использованием кинерадиографии (Herrel et al. 1996a, 1997, 1999b; Herrel and De Vree, 1999) показывает, что только гекконы и Cordylus имеют истинно связанный кинезис, при котором стрептостили (i.е. вращение квадратной кости в квадрато-чешуйчатом суставе) связано с мезокинезом во время кормления. Среди видов, включенных в настоящее исследование, змеи — единственная другая группа, которая демонстрирует поразительную степень внутричерепной подвижности. Представители других групп, таких как ангиды ( Gherronotus liocephalus , Ophisaurus apoda ) и вараниды ( Varanus ornatus , Varanus niloticus ), у которых в литературе сообщалось, что кинетические черепа не показывали движения на рентгеновских снимках. на мезокинетической оси во время кормления. Чтобы проверить потенциальное влияние уменьшения силы укуса на эффективность пищевого поведения у ящериц с кинетическим черепом, мы количественно оценили продолжительность внутриротовых транспортных циклов у ряда ящериц (Таблица 2). Анализ ковариации продолжительности транспортных циклов (длина черепа как ковариата) и тестирование различий среди животных с кинетическим черепом (гекконы, Cordylus и Nerodia ) и всех других показывают, что транспортировка значительно дольше у чешуек с кинетическим черепом. черепа (акинетические: 0.67 ± 0,37 с; кинетическая: 3,44 ± 3,27; F 2,17 = 13,77; P <0,01; Таблица 2, Рис.5). Таким образом, следствием кинетического черепа является увеличение продолжительности транспортных циклов. Количество циклов, необходимых для уменьшения объекта добычи, также больше (10,91 ± 5,44 против 31,92 ± 9,43; F 1,10 = 11,78; P <0,01) у ящериц с кинетическими черепами, и это означает, что эти чешуйчатым требуется значительно (почти в 10 раз) больше времени для транспортировки добычи.Это может снова иметь серьезные экологические последствия, поскольку это повысит подверженность потенциальным хищникам. Однако изменения в поведении, включая крипсис (например, Cordylus), , ночной образ жизни (гекконы) или нечастое кормление (змеи), могут устранить эти потенциальные недостатки. Рис. 5 Продолжительность транспортных циклов добычи у чешуек. Обратите внимание на то, что продолжительность внутриротовых транспортных циклов больше у чешуек с кинетическими черепами (гекконы, змеи и Cordylus ).Белые круги, гекконы; светло-серый круг, змеи; темно-серый, кордилис; черные, прочие ящерицы. Рис. 5 Продолжительность транспортных циклов добычи у чешуек. Обратите внимание на то, что продолжительность внутриротовых транспортных циклов больше у чешуек с кинетическими черепами (гекконы, змеи и Cordylus ). Белые круги, гекконы; светло-серый круг, змеи; темно-серый, кордилис; черные, прочие ящерицы. Хотя кажется, что краниальный кинезис является неблагоприятным побочным продуктом слабо построенного черепа, некоторые преимущества краниального кинезиса были предложены в литературе.Например, было высказано предположение, что кинетический механизм дополнительного сустава позволит быстрее открывать и закрывать челюсти у гекконов (Herrel et al. 2000). Однако это никогда не проверялось количественно. Мы проверили эту гипотезу, используя ограниченный набор данных по кинематическим фазам цикла зазоров. ANCOVA с длиной черепа как ковариант и тестирование различий в продолжительности быстрого открытия (FO) между животными с кинетическими черепами (например, гекконы и Cordylus ; змеи явно имеют совершенно другой кинетический механизм и не ожидается, что они будут быстрее) и с акинетическим черепом, однако, не имеет значения ( F 1,8 = 2.92; P = 0,13; Таблица 2). Тем не менее, если только гекконы кодируются как кинетические, различия в продолжительности ОС очень значительны ( F 1,8 = 14,89; P <0,01). Таким образом, протестированные здесь гекконы отличаются от кордилид тем, что имеют гораздо более быструю фазу FO (Таблица 2, Рис. 6A). Хотя поначалу это довольно удивительно, наши морфологические данные позволяют предположить, почему это может быть так. Хотя чешуйки с кинетическими черепами (гекконы, Cordylus и змеи) имеют значительно большую MPPt для данной длины черепа, чем чешуйчатые с акинетическими черепами (анализ моделирования на основе морфологического дерева: F тип = 7.74; молекулярное дерево: F фил = 10,45; F trad = 11,68, P <0,05; Таблица 1, Рис. 7), эта закономерность в значительной степени обусловлена непропорционально большим MPPt у змей и гекконов (Рис. 7). MPPt участвует в удлинении черепной части у гекконов и может способствовать более быстрому вращению челюстей у этих животных. Cordylus , с другой стороны, имеет относительно меньшую MPPt и, следовательно, не может вращать свою верхнюю челюсть так быстро, как гекконы.Очевидно, что для окончательной проверки закономерностей, предлагаемых этими данными, необходимо больше данных. Рис. 6 ( A ) Продолжительность фазы быстрого раскрытия челюстей во время транспортировки добычи. Обратите внимание, насколько гекконы очень быстрые. С другой стороны, Cordylus , несмотря на кинетический череп, не быстрее, чем ящерицы с акинетическим черепом. ( B) продолжительность фазы быстрого смыкания челюстей во время транспортировки добычи.Обратите внимание на то, как гекконы (белые круги) снова очень быстрые. Cordylus (серый круг) также быстрее, чем ящерицы с акинетическими черепами (черный круг). Рис. 6 ( A ) Продолжительность фазы быстрого раскрытия челюстей во время транспортировки добычи. Обратите внимание, насколько гекконы очень быстрые. С другой стороны, Cordylus , несмотря на кинетический череп, не быстрее, чем ящерицы с акинетическим черепом. ( B) продолжительность фазы быстрого смыкания челюстей во время транспортировки добычи.Обратите внимание на то, как гекконы (белые круги) снова очень быстрые. Cordylus (серый круг) также быстрее, чем ящерицы с акинетическими черепами (черный круг). Рис. 7 График, изображающий взаимосвязь между длиной черепа и массой MPPt, участвующего в вытягивании морды во время раскрытия челюсти у гекконов. Обратите внимание на то, что у гекконов большие мышцы MPPt, что может объяснить чрезвычайно быструю фазу раскрытия. Cordylus , несмотря на кинетический череп, не имеет MPPt больше, чем у ящериц с акинетическим черепом. Рис. 7 График, изображающий взаимосвязь между длиной черепа и массой MPPt, участвующего в вытягивании морды во время раскрытия челюсти у гекконов. Обратите внимание на то, что у гекконов большие мышцы MPPt, что может объяснить чрезвычайно быструю фазу раскрытия. Cordylus , несмотря на кинетический череп, не имеет MPPt больше, чем у ящериц с акинетическим черепом. Во время закрытия челюсти, однако, приводящие челюсти отвечают как за вентрадное вращение морды, так и за подъем нижней челюсти, поэтому ожидается, что гекконы и Cordylus будут быстрее, чем ящерицы с акинетическими черепами.Действительно, различия в продолжительности ФК значительно различаются между видами с кинетическим и акинетическим черепами ( F 1,8 = 11,66; P <0,01). Таким образом, гекконы и кордилиды, включенные в настоящий анализ, схожи в том, что имеют короткую продолжительность FC (таблица 2, рис. 6B). Быстрые движения смыкания челюстей у ящериц с кинетическим черепом могут быть важны, поскольку они могут дать потенциальное преимущество при питании неуловимой, подвижной добычей. Хотя это никогда ранее не оценивалось количественно, это должно предоставить интересную возможность для дальнейших исследований. В заключение, наши данные позволяют предположить, что конструкция черепа у ящериц определяется ограничениями минимального дизайна, связанными с укусом. Конструктивные требования, налагаемые другими факторами селективного давления (например, ночной жизнью), могут конкурировать с функциональными требованиями к прочности черепа, что может иметь далеко идущие последствия для использования системы кормления и, в конечном итоге, для экологии животных. Таким образом, черепная система ящерицы действительно кажется идеальной системой для изучения того, как различные селективные давления и потенциальные ограничения могут влиять на формирование эволюции сложной интегрированной системы. БлагодарностиАвторы хотели бы поблагодарить Тима Хайэма и Питера Уэйнрайта за приглашение представить эту статью на симпозиуме по эволюции пищевого поведения позвоночных на ежегодном собрании SICB в 2007 г. и SICB за финансовую поддержку. Мы также хотели бы поблагодарить импортеров животных в Бельгии (Anaconda Reptiles, Fantasia Reptiles) и Нидерландах (торговый дом Chameleon) за предоставление умерших образцов для морфологического анализа. А.Х. хотел бы поблагодарить Journal of Experimental Biology за предоставленный грант на поездку, который позволил ему посетить симпозиум 2001 г. по специализированным мышцам, а затем провести полевые работы в Австралии. Полевые работы в Австралии проводились по разрешению E24474-1. А.Х. является докторантом Фонда научных исследований, Фландрия, Бельгия (FWO-Vl). Исследование финансируется грантом на докторскую степень Института содействия инновациям через науку и технологии во Фландрии (IWT-Vlaanderen) В.S., грант Бельгийско-американского образовательного фонда К.М.и грант на проезд журнала экспериментальной биологии J.M. Ссылки.Краниальный кинезис у ящериц: разновидности, использование и происхождение. В: Hecht M, Macintyre R, Clegg M, editors ,Evolutionary biology ,1998 , vol.30 New York Plenum Press (стр.323 —57 ),,,.Экологические наблюдения за ящерицей Xenosaurus grandis в Куаутлапан, Веракрус, Мексика ,Biotropica ,1995 , vol.27 (стр.18 —32 ).К строительной морфологии рыб-чихлид (Teleostei, Perciformes) ,Neth J Zool ,1983 , vol.33 (стр.357 —424 ),,.Биомеханика подъязычной системы чешуи. В: Белс В.Л., Шардон М., Вандевалле П., редакторы ,Биомеханика питания позвоночных ,1994 Берлин Springer Verlag (стр.197 —240 ),,.Не полностью защищенные убежища: селекция и связанные с ней защиты ящерицей, Cordylus cordylus (Squamata: Cordylidae) ,Ethology ,1999 , vol.105 (стр.687 —700 ),.Кинематика пищевого поведения у Oplurus cuvieri (Reptilia: Iguanidae) ,J Exp Biol ,1992 , vol.170 (стр.155 —186 ),.Влияние ошибок длины ветвей на выполнение филогенетически независимых контрастов ,Syst Biol ,1998 , vol.47 (стр.654 —672 ),,.Филогенетические отношения внутри чешуйчатых. В: Estes R, Pregill G, редакторы ,Филогенетические отношения семейств ящериц ,1988 Stanford Stanford University Press (стр.119 —281 ).Филогении и сравнительный метод ,Am Nat ,1985 , vol.125 (стр.1 —15 ) и др.Ранняя эволюция ядовитой системы ящериц и змей ,Nature ,2005 , vol.439 (стр.584 —8 ).Механизм питания змей и его возможная эволюция ,Am Zool ,1961 , vol.1 (стр.217 —27 ),,,.Филогенетический анализ ковариации с помощью компьютерного моделирования ,Syst Biol ,1993 , vol.42 (стр.265 —92 ),,.Введение в филогенетические статистические методы с новым методом для доверительных интервалов для предковых состояний ,Am Zool ,1999 , vol.39 (стр.374 —88 ),,.Скваматические отношения, основанные на последовательностях ядерной ДНК C-mos: увеличение выборки таксонов улучшает поддержку начальной загрузки ,Amphibia Reptilia ,2001 , vol.22 (стр.235 —42 ),.Кинематика внутриротового транспорта и глотания у травоядных ящериц Uromastix acanthinurus ,J Exp Biol ,1999 , vol.202 (стр.1127 —37 ),,.Кинематика питания ящерицы Agama stellio ,J Exp Biol ,1996 , vol.199 (стр.1727 —42 ),,.Половой диморфизм размера головы у Podarcis hispanica atrata : Проверка гипотезы диетической дивергенции с помощью анализа силы укуса ,Neth J Zool ,1996 , vol.46 (стр.253 —62 ),,,.Механика овофагии у бисерной ящерицы ( Heloderma horridum ) ,J Herpetol ,1997 , vol.31 (стр.383 —93 ),,.Экоморфология устройства для кормления ящериц: подход к моделированию ,Neth J Zool ,1998 , vol.48 (стр.1 —25 ),,,.Половой диморфизм размера головы у Gallotia galloti ; проверка гипотезы дивергенции ниш с помощью функционального анализа ,Funct Ecol ,1999 , vol.13 (стр.289 —97 ),,.Модуляторная сложность кормового репертуара у сцинтилевых ящериц ,J Comp Physiol A ,1999 , vol.184 (стр.501 —18 ),,,.Краниальный кинезис у ящериц гекконид ,J Exp Biol ,1999 , vol.202 (стр.3687 —98 ),,.Краниальный кинез у гекконов: функциональные последствия ,J Exp Biol ,2000 , vol.203 (стр.1415 —23 ),,,.Влияние характеристик укуса на диету у двух видов ящериц-чешуек ,Can J Zool ,2001 , vol.79 (стр.662 —70 ),,,.Эволюция двигательных паттернов кормления у ящериц: модуляционная сложность и ограничения ,Am Zool ,2001 , vol.41 (стр.1311 —20 ),,.Форма головы и характеристики укуса ящериц-ксенозавров ,J Exp Zool ,2001 , vol.290 (стр.101 —7 ),,,.Показатели поклевки у клариидных рыб с гипертрофированными аддукторами челюстей по данным моделирования прикуса ,J Morphol ,2002 , vol.253 (стр.196 —205 ),,.Эволюция укусов черепах ,J Evol Biol ,2002 , vol.15 (стр.1083 —94 ),,.Всеядность ящериц-лацертидов: адаптивная эволюция или ограничение? ,Дж Evol Biol ,2004 , т.17 (стр.974 —84 ),,,.Экологические последствия онтогенетических изменений формы головы и характеристик укуса ямайской ящерицы Anolis lineatopus ,Biol J Linn Soc ,2006 , vol.89 (стр.443 —54 ).Режим травоядности и кормодобывания ящериц. В: Reilly SM, McBrayer LM, Miles DB, редакторы ,Эволюционные последствия режима кормодобывания у ящериц Cambridge Cambridge University Press ,,,,.Морфология, продуктивность и боеспособность самцов ящериц, Gallotia galloti ,Funct Ecol ,2005 , vol.19 (стр.800 —7 ),,,,.Тактика ведения боя различается по двум разным мужским фенотипам у ящерицы: тяжелые и легкие схватки ,Proc R Soc Lond B ,2004 , vol.271 (стр.2501 —8 ),.Характеристики оружия, а не размер, определяют успех спаривания и потенциальную репродуктивную способность у ошейниковой ящерицы ( Crotaphytus collaris ) ,Am Nat ,2005 , vol.166 (стр.426 —36 ),,,,.Зазорчатые дисплеи показывают и усиливают механический индекс эффективности оружия ,Am Nat ,2006 , vol.168 (стр.100 —13 ).Конвергентная эволюция и корреляция характеров у роющих рептилий: к разрешению плоских взаимоотношений ,Biol J Linn Soc ,1998 , vol.65 (стр.369 —453 ),,.Естественная история мексиканской чешуйчатой ящерицы, Xenosaurus rectocollaris ,Herp Nat Hist ,1996 , vol.4 (стр.151 —4 ),.Филогенетический анализ коррелированной эволюции непрерывных признаков: исследование с помощью моделирования ,Evolution ,1991 , vol.45 (стр.534 —57 ).Краниальный кинезис у лепидозавров: черепа в движении. В: Aerts P, D’Aout K, Herrel A, Van Damme R, редакторы ,Темы функциональной и экологической морфологии позвоночных ,2002 Maastricht Shaker Publishing (стр.15 —46 ),.Корреляция между формой черепа ящерицы и диетой: количественный филогенетический анализ ,Biol J Linn Soc ,2005 , vol.86 (стр.433 —66 ),.Кинематика отлова ящериц, питающихся муравьями ,J Exp Biol ,2005 , vol.208 (стр.113 —27 ).Ранняя потеря и множественное возвращение нижней височной аркады у диапсидных рептилий ,Naturwissenschaften ,2003 , vol.90 (стр.473 —6 ),.Утрата нижней височной аркады у диапсидных рептилий ,Zool J Linn Soc ,1981 , vol.72 (стр.203 —17 ).Кормление лепидозавров ,Кормление ,2000 Сан-Диего Academic Press (стр.175 —291 ),,,.Молекулярная филогенетика чешуйчатых: положение змей, амфисбенов и дибамид и корень чешуйчатого дерева ,Syst Biol ,2004 , vol.53 (стр.735 —57 ),,.Взаимосвязь между размером головы, силой укуса, эффективностью обращения с добычей и рационом у двух симпатрических лацертидных ящериц ,Funct Ecol ,2002 , vol.16 (стр.842 —50 ),.Молекулярные доказательства наземного происхождения змей ,Proc R Soc Lond B ,2004 , vol.271 (стр.S226 —9 ),,,.Функциональное значение «размера жертвы» у водных змей ( Nerodia fasciata , Colubridae) ,Oecologia ,2006 , vol.147 (стр.204 —11 ),.Размер глаз у гекконов: асимметрия, аллометрия, половой диморфизм и поведенческие корреляты ,J Morphol ,2006 , vol.267 (стр.1486 —500 )Заметки автора© Автор 2007.Опубликовано Oxford University Press от имени Общества интегративной и сравнительной биологии. Все права защищены. За разрешениями обращайтесь по электронной почте: journals.permissions@oxfordjournals.org. .Оставить комментарий
|